Elastin Anticorps
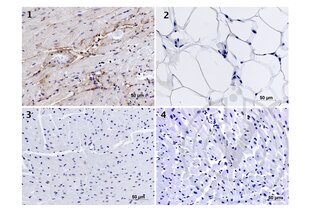
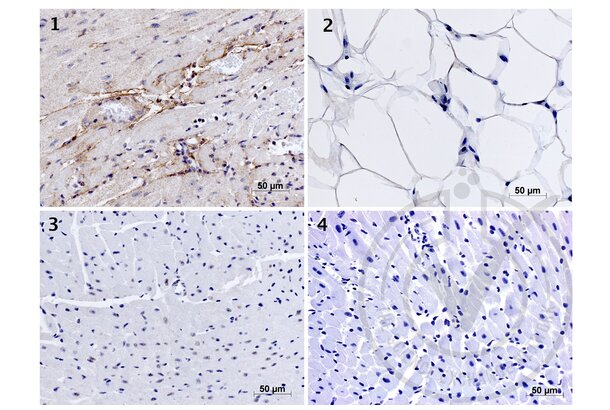 Elastin anticorps (AA 681-786) (ABIN734003)
Elastin anticorps (AA 681-786) (ABIN734003)
ELN Reactivité: Humain, Souris, Rat WB, ELISA, IHC (p), IHC (fro), IF (cc), IF (p), FACS Hôte: Lapin Polyclonal unconjugated
ELN Reactivité: Humain WB, ELISA, ICC, IHC (p), IHC (fro) Hôte: Lapin Polyclonal unconjugated
ELN Reactivité: Souris WB, IHC, IP, ICC Hôte: Lapin Polyclonal unconjugated
Elastin Anticorps par Réactivité
Trouvez Elastin Anticorps pour une variété d'espèces telles que anti-Human Elastin, anti-Mouse Elastin, anti-Rat Elastin. Les espèces listées ci-dessous sont parmi celles disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
Elastin Anticorps par Hote
On trouve ici des Elastin Anticorps avec un Hote spécifique. Les Hote mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
Elastin Anticorps par Epitope
Trouver Elastin Anticorps avec un épitope spécifique. Les épitopes listés ci-dessous sont parmi ceux disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
Elastin Anticorps par Clonalité
Trouvez les Elastin Anticorps monoclonaux ou polyclonaux disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
Elastin Anticorps par Clone
On trouve ici des Elastin Anticorps avec un Clone spécifique. Les Clone mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
Elastin Anticorps par Conjugué
Trouvez Elastin Anticorps avec un conjugué spécifique tel que Biotin, FITC, Alexa Fluor 488. Les conjugués listés ci-dessous sont parmi ceux disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
Elastin Anticorps fréquemment utilisés
- (4)
- (7)
- (1)
- (8)
- (3)
- (3)
- (3)
- (3)
- (3)
- (2)
- (2)
- (2)
- (1)
- (1)
- (1)
- (1)
- (2)
Dernières publications pour nos Elastin Anticorps
: "Tropoelastin: A novel marker for plaque progression and instability." dans: Circulation. Cardiovascular imaging, Vol. 11, Issue 8, (2018) (PubMed).: "Vasoprotective effects of urocortin 1 against atherosclerosis in vitro and in vivo." dans: PLoS ONE, Vol. 9, Issue 12, pp. e110866, (2014) (PubMed).
: "Stimulatory effects of cardiotrophin 1 on atherosclerosis." dans: Hypertension, Vol. 62, Issue 5, pp. 942-50, (2013) (PubMed).
: "Elastogenesis at the onset of human cardiac valve development." dans: Development (Cambridge, England), Vol. 140, Issue 11, pp. 2345-53, (2013) (PubMed).
: "Immunofluorescence and fluorescent-protein tagging show high correlation for protein localization in mammalian cells." dans: Nature methods, Vol. 10, Issue 4, pp. 315-23, (2013) (PubMed).
: "Piezoelectric PU/PVDF electrospun scaffolds for wound healing applications." dans: Colloids and surfaces. B, Biointerfaces, Vol. 96, pp. 29-36, (2012) (PubMed).
: "The influence of elastic components of the venous wall on the biomechanical properties of different veins used for arterial reconstruction." dans: European journal of vascular and endovascular surgery : the official journal of the European Society for Vascular Surgery, Vol. 40, Issue 2, pp. 224-9, (2010) (PubMed).
: "Impact of cryopreservation on extracellular matrix structures of heart valve leaflets." dans: The Annals of thoracic surgery, Vol. 81, Issue 3, pp. 918-26, (2006) (PubMed).
: "Connection between elastin haploinsufficiency and increased cell proliferation in patients with supravalvular aortic stenosis and Williams-Beuren syndrome." dans: American journal of human genetics, Vol. 71, Issue 1, pp. 30-44, (2002) (PubMed).
: "Characterization of biologically active domains on elastin: identification of a monoclonal antibody to a cell recognition site." dans: Biochemistry, Vol. 25, Issue 18, pp. 5172-6, (1986) (PubMed).
Pseudonymes pour Elastin Anticorps
elastin (ELN) Anticorpselastin (Eln) Anticorps
elastin (LOC574132) Anticorps
elastin (Tcur_2481) Anticorps
elastin (Kfla_2289) Anticorps
elastin (LOC100620140) Anticorps
AI385707 Anticorps
AI480567 Anticorps
E030024M20Rik Anticorps
ELN Anticorps
RATTREL11 Anticorps
SVAS Anticorps
TREL11 Anticorps
Trela Anticorps
Trela26 Anticorps
WBS Anticorps
WS Anticorps
Avez-vous cherché autre chose?
- Elastase 3B Anticorps
- Elastase Anticorps
- ELAPOR1 Anticorps
- ELANE Anticorps
- ELAC2 Anticorps
- ELAC1 Anticorps
- EIF6 Anticorps
- EIF5B Anticorps
- EIF5AL1 Anticorps
- EIF5A2 Anticorps
- EIF5 Anticorps
- EIF4H Anticorps
- EIF4G3 Anticorps
- EIF4G2 Anticorps
- EIF4G1 Anticorps
- EIF4ENIF1 Anticorps
- EIF4EBP3 Anticorps
- eIF4EBP2 Anticorps
- eIF4EBP1 Anticorps
- EIF4E2 Anticorps
- ELAV (Embryonic Lethal, Abnormal Vision, Drosophila)-Like 1 (Hu Antigen R) Anticorps
- ELAVL2 Anticorps
- ELAVL4 Anticorps
- ELF1 Anticorps
- ELF2 Anticorps
- ELF3 Anticorps
- Elf4 Anticorps
- ELF5 Anticorps
- ELFN1 Anticorps
- ELFN2 Anticorps
- ELK1 Anticorps
- ELK3 Anticorps
- ELK4 Anticorps
- ELL Anticorps
- ELL2 Anticorps
- ELL3 Anticorps
- ELMO1 Anticorps
- ELMO2 Anticorps
- ELMO3 Anticorps
- ELMOD1 Anticorps



