C9 Anticorps
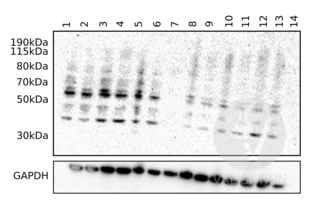
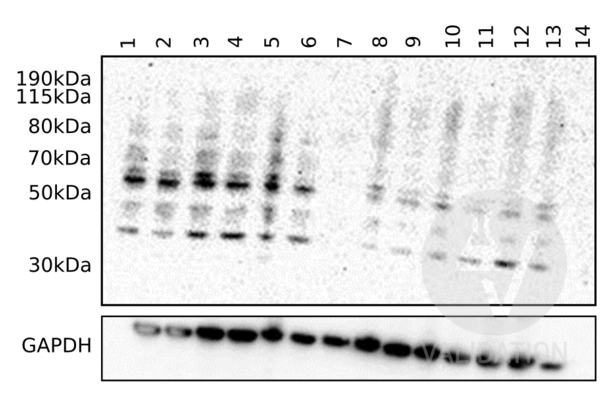 C9 anticorps (Center) (ABIN657704)
C9 anticorps (Center) (ABIN657704)
C9 Reactivité: Humain WB, IHC (p) Hôte: Lapin Polyclonal RB33918 unconjugated
C9 Reactivité: Humain WB, IHC, IP, ICC Hôte: Lapin Polyclonal unconjugated
C9 Reactivité: Souris WB, IHC, IP, ICC Hôte: Lapin Polyclonal unconjugated
C9 Anticorps par Grade
On trouve ici des C9 Anticorps avec un Grade spécifique. Les Grade mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
C9 Anticorps par Réactivité
Trouvez C9 Anticorps pour une variété d'espèces telles que anti-Human C9, anti-Mouse C9, anti-Horse C9. Les espèces listées ci-dessous sont parmi celles disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
C9 Anticorps par Hote
On trouve ici des C9 Anticorps avec un Hote spécifique. Les Hote mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
C9 Anticorps par Clonalité
Trouvez les C9 Anticorps monoclonaux ou polyclonaux disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
C9 Anticorps par Clone
On trouve ici des C9 Anticorps avec un Clone spécifique. Les Clone mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
C9 Anticorps fréquemment utilisés
- (3)
- (1)
- (4)
- (3)
- (2)
- (3)
- (1)
- (1)
- (2)
Dernières publications pour nos C9 Anticorps
: "Diabetes-related changes in the protein composition and the biomechanical properties of human retinal vascular basement membranes." dans: PLoS ONE, Vol. 12, Issue 12, pp. e0189857, (2018) (PubMed).: "Angiogenic factor imbalance precedes complement deposition in placentae of the BPH/5 model of preeclampsia." dans: FASEB journal : official publication of the Federation of American Societies for Experimental Biology, Vol. 32, Issue 5, pp. 2574-2586, (2018) (PubMed).
: "Target deletion of complement component 9 attenuates antibody-mediated hemolysis and lipopolysaccharide (LPS)-induced acute shock in mice." dans: Scientific reports, Vol. 6, pp. 30239, (2016) (PubMed).
: "Selectivity analysis of single binder assays used in plasma protein profiling." dans: Proteomics, Vol. 13, Issue 23-24, pp. 3406-10, (2013) (PubMed).
: "Defining the CD59-C9 binding interaction." dans: The Journal of biological chemistry, Vol. 281, Issue 37, pp. 27398-404, (2006) (PubMed).
: "The human complement C9 gene: structural analysis of the 5' gene region and genetic polymorphism studies." dans: European journal of immunogenetics : official journal of the British Society for Histocompatibility and Immunogenetics, Vol. 28, Issue 5, pp. 515-22, (2002) (PubMed).
: "Induction of complement C9 messenger RNAs in human neuronal cells by inflammatory stimuli: relevance to neurodegenerative disorders." dans: Experimental gerontology, Vol. 36, Issue 7, pp. 1179-88, (2001) (PubMed).
: "Analysis of C5b-8 binding sites in the C9 molecule using monoclonal antibodies: participation of two separate epitopes of C9 in C5b-8 binding." dans: Molecular immunology, Vol. 29, Issue 7-8, pp. 911-6, (1992) (PubMed).
Pseudonymes pour C9 Anticorps
complement C9 (C9) Anticorpscomplement C9 (c9) Anticorps
complement component 9 (C9) Anticorps
complement component 9 L homeolog (c9.L) Anticorps
complement component 9 (c9) Anticorps
c9 Anticorps
LOC100037951 Anticorps
LOC100136130 Anticorps
wu:fb60b05 Anticorps
wu:fd50e04 Anticorps
zgc:112272 Anticorps
Avez-vous cherché autre chose?
- C8orf80 Anticorps
- C8orf55 Anticorps
- C8orf4 Anticorps
- C8ORF37 Anticorps
- C8G Anticorps
- C8B Anticorps
- C8A Anticorps
- C7orf64 Anticorps
- C7orf49 Anticorps
- C7orf30 Anticorps
- C7orf10 Anticorps
- C7 Anticorps
- C6orf89 Anticorps
- C6orf25 Anticorps
- C6orf204 Anticorps
- C6orf192 Anticorps
- C6orf168 Anticorps
- C6orf150 Anticorps
- C6ORF136 Anticorps
- C6orf130 Anticorps
- C9orf100 Anticorps
- C9orf11 Anticorps
- C9orf156 Anticorps
- C9orf21 Anticorps
- C9orf24 Anticorps
- C9orf25 Anticorps
- C9orf3 Anticorps
- C9orf6 Anticorps
- C9ORF72 Anticorps
- C9orf86 Anticorps
- C9orf89 Anticorps
- CA 19-9 Anticorps
- CA1 Anticorps
- CA10 Anticorps
- CA11 Anticorps
- CA12 Anticorps
- CA13 Anticorps
- CA14 Anticorps
- CA2 Anticorps
- CA3 Anticorps



