HYAL1 Anticorps
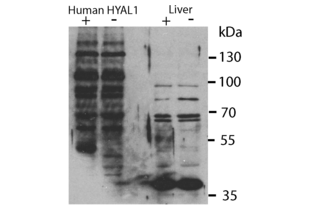
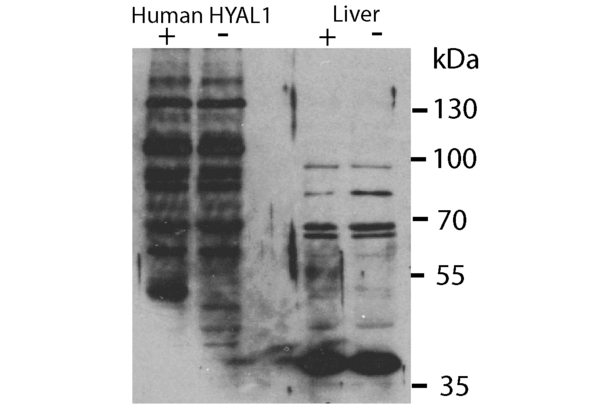 HYAL1 anticorps (N-Term) (ABIN2784342)
HYAL1 anticorps (N-Term) (ABIN2784342)
HYAL1 Reactivité: Humain, Souris, Rat, Boeuf (Vache), Chien, Cobaye, Cheval, Lapin WB, IHC Hôte: Lapin Polyclonal unconjugated
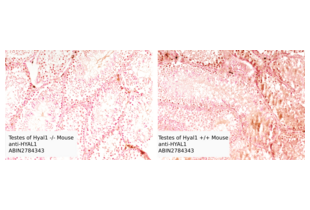
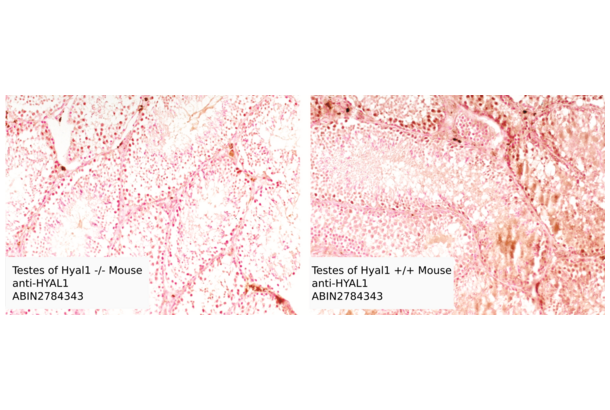 HYAL1 anticorps (N-Term) (ABIN2784343)
HYAL1 anticorps (N-Term) (ABIN2784343)
HYAL1 Reactivité: Humain, Souris, Rat, Boeuf (Vache), Chien, Cobaye, Cheval, Lapin WB, IHC Hôte: Lapin Polyclonal unconjugated
HYAL1 Reactivité: Humain, Souris, Rat WB, IHC Hôte: Lapin Polyclonal unconjugated
HYAL1 Anticorps par Grade
On trouve ici des HYAL1 Anticorps avec un Grade spécifique. Les Grade mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
HYAL1 Anticorps par Hote
On trouve ici des HYAL1 Anticorps avec un Hote spécifique. Les Hote mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
HYAL1 Anticorps par Clonalité
Trouvez les HYAL1 Anticorps monoclonaux ou polyclonaux disponibles. Cliquez sur un lien pour accéder aux produits correspondants.
HYAL1 Anticorps par Clone
On trouve ici des HYAL1 Anticorps avec un Clone spécifique. Les Clone mentionnés ici sont quelques-uns de ceux qui sont disponibles. Un clic sur le lien correspondant permet d'accéder aux produits
HYAL1 Anticorps fréquemment utilisés
- (5)
- (1)
- (2)
- (1)
- (4)
- (4)
- (5)
- (3)
- (1)
- (2)
- (2)
- (2)
- (1)
- (1)
- (2)
- (2)
- (3)
- (1)
- (1)
Dernières publications pour nos HYAL1 Anticorps
: "Hyaluronan synthases and hyaluronidases in nasal polyps." dans: European archives of oto-rhino-laryngology : official journal of the European Federation of Oto-Rhino-Laryngological Societies (EUFOS) : affiliated with the German Society for Oto-Rhino-Laryngology - Head and Neck Surgery, Vol. 273, Issue 7, pp. 1801-8, (2016) (PubMed).: "Evaluation of protein biomarkers of prostate cancer aggressiveness." dans: BMC cancer, Vol. 14, pp. 244, (2014) (PubMed).
: "Inverse expression of hyaluronidase 2 and hyaluronan synthases 1-3 is associated with reduced hyaluronan content in malignant cutaneous melanoma." dans: BMC cancer, Vol. 13, pp. 181, (2013) (PubMed).
: "Increased hyaluronic acid content in idiopathic pulmonary arterial hypertension." dans: The European respiratory journal, Vol. 32, Issue 6, pp. 1504-12, (2008) (PubMed).
: "Neurofibromatosis-1 (Nf1) heterozygous brain microglia elaborate paracrine factors that promote Nf1-deficient astrocyte and glioma growth." dans: Human molecular genetics, Vol. 16, Issue 9, pp. 1098-112, (2007) (PubMed).
Pseudonymes pour HYAL1 Anticorps
hyaluronoglucosaminidase 1 (HYAL1) Anticorpshyaluronoglucosaminidase 1 (Hyal1) Anticorps
Hya1 Anticorps
HYAL-1 Anticorps
Hyal-1 Anticorps
LUCA1 Anticorps
NAT6 Anticorps
Avez-vous cherché autre chose?
- HWP1 Anticorps
- HVEM Anticorps
- HVD3 Anticorps
- HVCN1 Anticorps
- HUWE1 Anticorps
- HUS1B Anticorps
- HUS1 Anticorps
- Huntingtin Anticorps
- HuC/ELAVL3 Anticorps
- HTRA4 Anticorps
- HTRA3 Anticorps
- HTRA2 Anticorps
- HTRA1 Anticorps
- HTR7 Anticorps
- HTR5B Anticorps
- HTR3E Anticorps
- HTR3D Anticorps
- HTR2C Anticorps
- HTR2A Anticorps
- HTR1F Anticorps
- HYAL2 Anticorps
- HYAL3 Anticorps
- HYAL4 Anticorps
- Hyaluronic Acid Anticorps
- Hydrocephalus Inducing Anticorps
- Hydroxyacid Oxidase 2 Anticorps
- Hydroxyproline Anticorps
- Hydroxyprostaglandin Dehydrogenase 15-(NAD) Anticorps
- Hydroxysteroid (11-Beta) Dehydrogenase 1-Like Anticorps
- HYI Anticorps
- HYLS1 Anticorps
- HYOU1 Anticorps
- Hypoxanthine phosphoribosyltransferase 1 Anticorps
- IAH1 Anticorps
- IARS Anticorps
- IB2 Anticorps
- IBA57 Anticorps
- IBSP Anticorps
- IBTK Anticorps
- ICA1 Anticorps



