CTNNB1 Anticorps
Choisissez l’un des 247 anticorps CTNNB1 de notre liste de produits basés sur 68 publications et 314 images de validation.
Découvrez des anticorps CTNNB1 fiables issus d’une large sélection de fabricants renommés. Notre portefeuille permet une détection précise de CTNNB1 dans plusieurs espèces, y compris Human, Mouse, Rat, Dog, Monkey, Chicken, Cow, Zebrafish (Danio rerio), Pig, Horse, Rabbit, Xenopus laevis, Hamster, Bat, Guinea Pig, Sheep, Opossum, et prend en charge diverses applications de recherche telles que WB, IHC, ELISA, IF, FACS.
Trouvez rapidement l’anticorps adapté grâce à nos outils de recherche, de filtrage et de comparaison. Chaque page produit fournit des données de validation détaillées, des références bibliographiques et des retours d’utilisateurs. Contrairement aux ressources dédiées uniquement à la comparaison, vous pouvez acheter directement les anticorps sur notre site. Choisissez parmi des anticorps polyclonaux, monoclonaux ou recombinants CTNNB1, issus de différents hôtes tels que Rabbit, Mouse, Alpaca. Parcourez nos produits ci-dessous et commandez directement pour soutenir vos recherches en toute confiance.
Les noms alternatifs de CTNNB1 sont CTNNB1, Ctnnb1, ctnnb1, ctnnb1.S, ctnnb1.L.
Filtrer les résultats:
247 Résultats
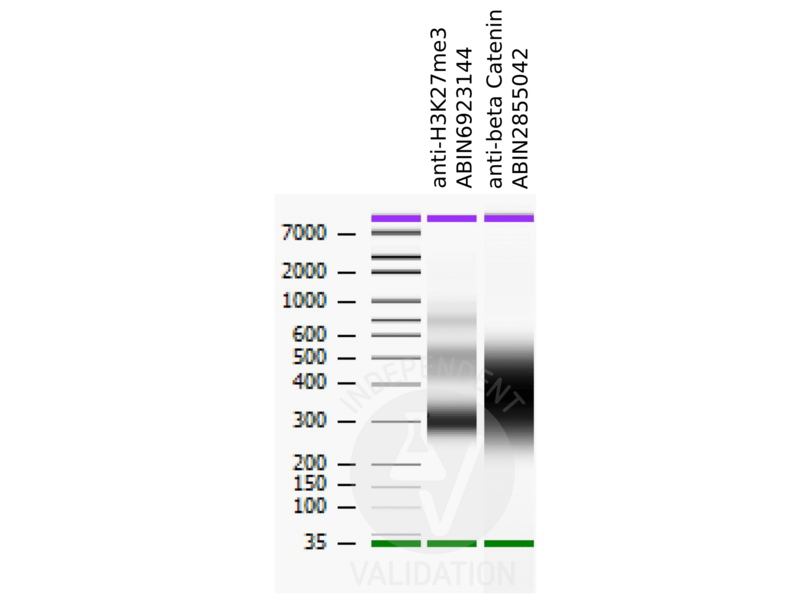
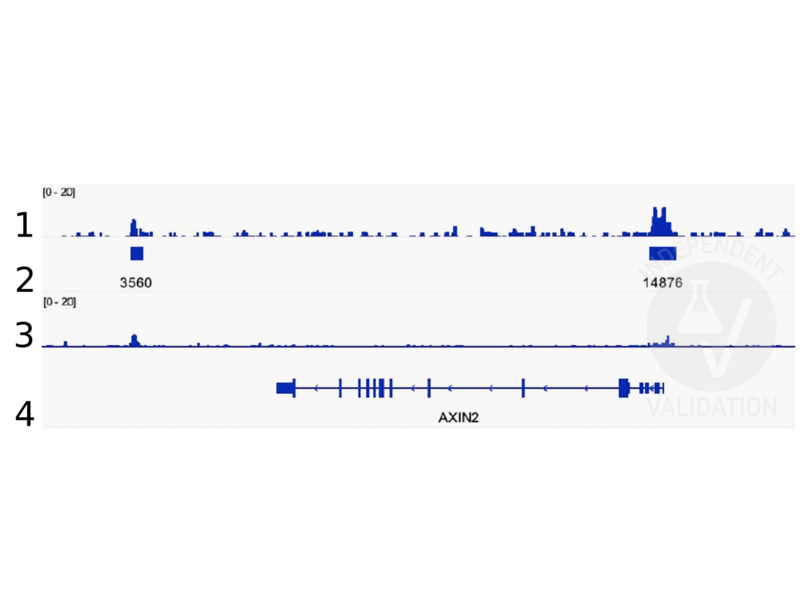
 11 références
11 références
 1 validation
5 références
6 références
5 références
1 reference
1 reference
1 reference
1 validation
5 références
6 références
5 références
1 reference
1 reference
1 reference
Dernières publications pour nos CTNNB1 Anticorps
: "Exhaustive identification of genome-wide binding events of transcriptional regulators." dans: Nucleic acids research, Vol. 52, Issue 7, pp. e40, (2024) (PubMed).
: "The β-catenin C terminus links Wnt and sphingosine-1-phosphate signaling pathways to promote vascular remodeling and atherosclerosis." dans: Science advances, Vol. 10, Issue 11, pp. eadg9278, (2024) (PubMed).
: "Acute Intoxication With Alcohol Reduces Trauma-Induced Proinflammatory Response and Barrier Breakdown in the Lung via the Wnt/β-Catenin Signaling Pathway." dans: Frontiers in immunology, Vol. 13, pp. 866925, (2022) (PubMed).
: "38" dans: , Vol. 1363, Issue Nucleic acids research, pp. 28-39, (1991)
: "A new cut&run low volume-urea (LoV-U) protocol optimized for transcriptional co-factors uncovers Wnt/b-catenin tissue-specific genomic targets." dans: Development (Cambridge, England), (2022) (PubMed).
: "Identification of candidate lncRNAs and circRNAs regulating WNT3/β-catenin signaling in essential hypertension." dans: Aging, Vol. 12, Issue 9, pp. 8261-8288, (2020) (PubMed).
: "A novel c-Kit/phospho-prohibitin axis enhances ovarian cancer stemness and chemoresistance via Notch3-PBX1 and β-catenin-ABCG2 signaling." dans: Journal of biomedical science, Vol. 27, Issue 1, pp. 42, (2020) (PubMed).
: "ARRDC3 suppresses colorectal cancer progression through destabilizing the oncoprotein YAP." dans: FEBS letters, Vol. 592, Issue 4, pp. 599-609, (2019) (PubMed).
: "Progranulin protects the mouse retina under hypoxic conditions via inhibition of the Toll‑like receptor‑4‑NADPH oxidase 4 signaling pathway." dans: Molecular medicine reports, Vol. 19, Issue 1, pp. 382-390, (2019) (PubMed).
Pseudonymes pour CTNNB1 Anticorps
catenin beta 1 (CTNNB1) Anticorpscatenin (cadherin associated protein), beta 1 (Ctnnb1) Anticorps
catenin beta 1 (Ctnnb1) Anticorps
catenin (cadherin-associated protein), beta 1 (ctnnb1) Anticorps
catenin beta 1 S homeolog (ctnnb1.S) Anticorps
catenin beta 1 L homeolog (ctnnb1.L) Anticorps
armadillo Anticorps
beta-catenin Anticorps
Bfc Anticorps
Catnb Anticorps
CHBCAT Anticorps
CTNNB Anticorps
ctnnb Anticorps
CTNNB1 Anticorps
ctnnb1 Anticorps
id:ibd2058 Anticorps
Jup Anticorps
LOC100217813 Anticorps
Mesc Anticorps
MRD19 Anticorps
wu:fb73e10 Anticorps
wu:fi81c06 Anticorps
wu:fk25h01 Anticorps
Avez-vous cherché autre chose?
- CTNNAL1 Anticorps
- CTNNA3 Anticorps
- CTNNA2 Anticorps
- CTNNA1 Anticorps
- CTLA4 Anticorps
- CTIF/KIAA0427 Anticorps
- CTHRC1 Anticorps
- CTH Anticorps
- CTGF Anticorps
- CTF18 Anticorps
- CTDSPL Anticorps
- CTDSP2 Anticorps
- CTDSP1 Anticorps
- CTDP1 Anticorps
- CTDNEP1A Anticorps
- CTD (Carboxy-Terminal Domain, RNA Polymerase II, Polypeptide A) Small Phosphatase Like 2 Anticorps
- CTCFL Anticorps
- CTCF Anticorps
- CTC1 Anticorps
- CTBS Anticorps
- CTNNBIP1 Anticorps
- CTNNBL1 Anticorps
- CTNND1 Anticorps
- CTNND2 Anticorps
- CTNS Anticorps
- CTP Synthase Anticorps
- CTPS2 Anticorps
- CTR9 Anticorps
- CTRB1 Anticorps
- CTRP5 Anticorps
- CTRP6 Anticorps
- CTRP7 Anticorps
- CTSA Anticorps
- CTSC Anticorps
- CTSO Anticorps
- CTTNBP2 Anticorps
- CTU2 Anticorps
- CTX-I Anticorps
- CUBN Anticorps
- CUE Domain Containing 1 Anticorps



